 Prof. Joseph Levine, M.D. is an emeritus associate professor in the Division of Psychiatry, Faculty of Health Sciences, Ben Gurion University in Israel. Prof. Levine is a certified psychiatrist with clinical experience in controlled trials of adult psychiatric disorders and in psychotherapy. He was awarded a NRSAD independent investigator grant for the study of Creatine Monohydrate in psychiatric disorders -- mainly Schizophrenia. He resides and treats patients in Tel Aviv and all of central Israel.
Prof. Joseph Levine, M.D. is an emeritus associate professor in the Division of Psychiatry, Faculty of Health Sciences, Ben Gurion University in Israel. Prof. Levine is a certified psychiatrist with clinical experience in controlled trials of adult psychiatric disorders and in psychotherapy. He was awarded a NRSAD independent investigator grant for the study of Creatine Monohydrate in psychiatric disorders -- mainly Schizophrenia. He resides and treats patients in Tel Aviv and all of central Israel.
Conversation 20: The evolution of the internalized characters
Greetings
Let's recall that we assumed that the core nucleus for the creation of internalized characters in humans are the areas that specialized in face recognition and especially the fusiform nucleus area.
This time we will ask what happens in a variety of animals from humans through other primates and vertebrates?
Do they also have internalized figures of their own species and perhaps of significant others [eg human faces in domesticated animals]?
In order to deal with this difficult question, we will try to go through studies that clarified the core of our hypothesis for the creation of internalized figures [if primary and diffuse and if complex] and that is the ability in higher animals to recognize their faces
We will raise the question of whether there are illustrations that reflect the processing of the most important facial data for an individual whether it is a person or an animal, those that the individual is more exposed to. Are these illustrations in which the processing is done in a better quality and maybe faster and with a higher resolution? Are these similar or different between humans and other primates and between humans and other vertebrates?
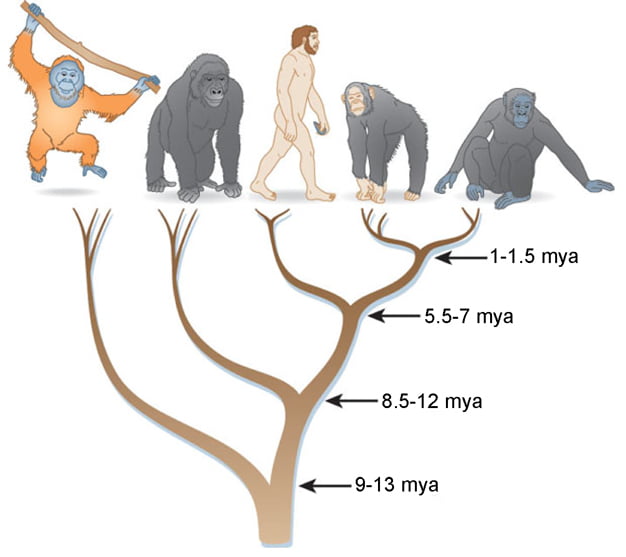
The upper group is the primate group
We will first discuss the important and comprehensive article by David Leopold and Gillian Rhodes who wrote about the comparative aspects of face perception between humans and other animal species.
David A. Leopold and Gillian Rhodes. A Comparative View of Face Perception. Journal of comparative psychology. (Washington, D.C. : 1983)

Gillian Rhodes – University of Western Australia
The authors point out that facial perception serves as the basis for a large part of the exchange of human social messages, and that a variety of information about a person can be extracted from one look at their face, including their identity, their emotional state and the direction of their attention. Neuropsychological experiments and fMRI [a kind of brain imaging that reflects brain activity] reveal a complex network of specialized areas in the human brain that support these face-reading skills. The authors examined the evolutionary roots of human face perception by investigating how different animal species see and respond to faces.They focused on behavioral experiments collected from both primates and non-primates, in which they examined the types of information that animals are able to extract from the faces of their peers, human experimenters and natural predators. These experiments revealed that faces are an important category of visual stimuli for animals in all major vertebrate types, which may reflect the early emergence of neural specialization for faces in vertebrate evolution. At the same time, certain aspects of face perception are only evident in primates and some other social mammals, and therefore may have evolved to suit the needs of complex social communication, since the human brain likely uses both primitive and recently evolved neural specializations for face processing. Comparative studies may therefore hold the key to understanding how these parallel circuits formed during human evolution.
For humans, they point out, faces are among the most important visual stimuli, a fact that becomes clear in social settings – since as a human species we constantly, almost obsessively, follow each other's faces, paying attention to fine details that can give us some insight into the emotional state , the level of involvement or the object of attention of our associates. Fast and high-quality processing of faces offers great social advantages, and allows a person to gather aspects and clues about the other's internal thought processes and to predict their behavior. But the question arises, how did humans come to put so much emphasis on this particular aspect of personal appearance? Is the ability to read faces a product of our society, well-tuned to meet the needs of human culture? Already in the nineteenth century, the authors claim that Darwin proposed that this is not the case, and that human facial expressions share much in common with those of many animals (Darwin, 1872). For Darwin, the study of the face was naturally categorized into its comparative and evolutionary context, a perspective adopted by only a small group of contemporary scholars.

Charles Darwin [1809-1882] and his book “Expression of Emotions in Man and Animals”.
The authors note that in their review they take a first step in the study of the evolution of face perception by reviewing a wide range of behavioral studies that provide insight into the following questions: (1) To what extent do faces constitute a “special” category of visual stimuli for nonhuman primates, as well as other mammals and vertebrates? That is, to what extent are faces important stimuli, evoking specific behavioral and neural responses? (2) What types of information are different animals able to extract from facial expressions?(3) How and when might different aspects of face perception emerge during evolution? Their review [they claim] covers behavioral experiments conducted in animals of many species related to visual recognition of the individual, predator detection, gaze tracking, and response to emotional expressions.
First they ask what types of information can be extracted from a face? Many studies have shown that face perception is multifaceted: not only do we recognize people, but we also track their faces to obtain a continuous stream of social information, from communicative gestures to emotional and attentive states. In our comparative review, we analyze separately different aspects of face perception, briefly described here for humans:
The recognition of single faces is in some ways the pinnacle of human visual performance. Because all faces have the same basic configural appearance (eg, two eyes above the nose and a mouth, sometimes called first-order configuration), people must be recognized by subtle deviations from this prototypical pattern, sometimes called second-order relational information or configuration. This form of recognition appears to rely on special brain regions that are selectively engaged by upright faces with normal contrast polarity.
Humans communicate their emotional state to others through a stereotypical posture of facial elements. Elaborate facial muscles contribute to a large repertoire of expressions that include the display of the teeth, the furrow of the brow and the closing of the eyes, some uniquely human and some not (Darwin, 1872). The appearance of an emotional expression can directly affect the emotional state of the observer. The look of the eyes serves as a particularly prominent emotional cue. It seems that some basic facial expressions are consistent among human populations, perhaps due to their adaptive value in evolution.
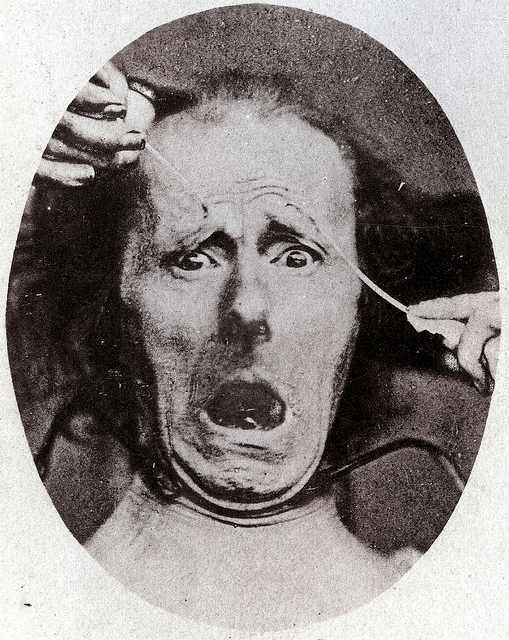

Photograph and illustration of a feeling of intense fear and anxiety from Charles Darwin's book [1872].
3) Look
In addition to its role in signaling emotional state, eye gaze also provides insight into a person's attentive state, including their level of engagement, intentions, or focus of interest. This aspect of face perception may be particularly developed in humans, as the sclera [simply put – the white part around the pupil] of the human eye is more visible than in other primates. Indeed, human infants primarily use the direction of the other's eyes, rather than the head, to determine the direction of the other person's gaze. Gaze reciprocity is a prominent feature of social interaction and is unusual in a number of psychiatric conditions, where patients typically avoid looking into the eyes of others.
Faces play an important role in human sexual attraction, which has been linked to average facial features, degree of symmetry and sexual dimorphism (masculinity in masculine faces, femininity in feminine faces).
The developmental trajectory of face perception is complex and only partially understood. Newborn infants recognize and visually orient to face-like patterns in preference to other complex patterns and form a preference for their mother's face within days of birth. They are also able to distinguish and imitate some basic facial expressions even in the hours after birth. During the first year of life, face processing mechanisms are "tuned" by repeated exposure to the faces of others and processing performance continues to improve throughout childhood and adolescence. The basis for this improvement remains controversial, with some claiming that sensitivity to second-order configuration [the ability to distinguish between individual faces] improves during childhood and others attribute it to the development of general-domain skills in attention and executive functions.
6) Neurological specialization
Different circuits in the brain support the perception of different types of information, a principle that is reflected both in brain-damaged patients and in imaging studies. The most prominent face-specialized area in the human brain is in the fusiform gyrus, where responses are stronger to faces than to any other category of stimuli. This region may receive basic perceptual representations of faces from a more occipital face-selective region and damage near these regions can lead to an inability to recognize individual faces. Some neurons in the medial temporal lobe are thought to represent people at a very general, abstract level, and respond strongly and selectively to faces. In contrast, the superior temporal sulcus appears to be sensitive to changing aspects of the face, as it responds selectively to emotional expression and eye gaze. The perception of eye gaze additionally activates the intraparietal sulcus, and its coding can be dissociated from that of identity and expression. Other neural structures respond to specific emotional expressions. For example, the amygdala selectively responds to fearful or unhappy facial expressions, while the orbitofrontal cortex responds to angry facial expressions.
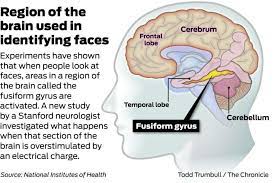
Thus human studies have shown that face perception involves a diverse set of skills supported by multiple specialized neural circuits and requires years to fully develop. Based on this framework, the authors turned to face perception by non-human species, and examined the extent to which different animals look at faces to extract different types of information, and addressed the developmental course and neural basis of face processing where the information is available. The authors asked to what extent faces are a special category of visual stimuli for nonhuman primates, other mammals, and other vertebrates. They review a wide range of experiments investigating the specific types of visual information different animals are unable to extract from faces. Then, based on the shared abilities of different animals, the authors tried to hypothesize the evolutionary history of different aspects of face perception, and how that history might be informative about our own face perception.
Non-human primates
Like humans, other primates are highly developed in their use of the face for social purposes, such as recognition, communication and mate selection. Primates have been extensively studied for how they perceive faces, mainly because of their evolutionary closeness to humans and the human-like ability to read faces, but also because they can be easily trained to respond to images and videos in a laboratory setting.
The development of facial processing in primates
The evolution of primate face processing is complex, and includes an innate tendency to look at the mother's face, along with a strong learning component. Chimpanzees can recognize their mother's face within a month of birth, with an increase in mutual gaze the following month. The subsequent exposure is also important for the development of their facial recognition skills, as evidenced by the fact that chimpanzees raised primarily among humans become experts with their human faces.
Similar to humans, infant monkeys prefer schematic faces over other stimuli, with spatial configuration being the important factor in the first month, and feature details becoming important later. Macaque monkeys who were deprived of any exposure to faces during the first 6-24 months of their lives showed an immediate interest in the faces that were presented afterwards, and acquired expertise with human or macaque faces thereafter depending on the experience they acquired. Thus, macaque monkeys exhibit an innate preference for faces, along with a window of plasticity [the ability to develop and change a particular brain capacity] (abnormally extended in this case) in which their perceptual skills for a given category of faces are sharpened. A similar developmental refinement has also been observed in humans, whose ability to distinguish monkey faces decreases during development, apparently due in part to widespread selective exposure to human faces.
In general, the authors note that primates extract a rich amount of social information from the faces of their peers. For the three species that have been studied in the most detail, humans, chimpanzees, and macaques, the many shared features of face processing, along with selective responses to seemingly similar faces in the brain, suggest that our last common ancestor, which lived more than 20 million years ago, may have enjoyed a similar situation in the context of face recognition. The functional and anatomical similarities raise the possibility of evolutionary homology (similarity due to common descent) between the primary face-selective cortical areas in macaque and man, albeit with a ventral shift in their anatomical location in humans. While neural specialization for faces has not been studied in New World monkeys, the few behavioral studies suggest that they also have sophisticated face perception, which may place the ancient origins of complex face processing as far back as purebred anthropoid primates, which lived more than 40 million years ago.
The authors ask whether the refined face perception abilities we share with chimpanzees and macaques evolved to fit the social needs of primates who form interactive relationships, or did primate sociability exploit existing state and abilities with faces?
Mammals
Compared to primates, other mammals rely less on sight and more on smell and hearing for social communication. However, many mammals have good visual acuity, although the use of vision in face perception has only been assessed in a few species. Curiously, the most studied non-primate species, which is sheep, shows a pronounced visual specialization for faces at both the behavioral and neural levels. In the mid-1980s, Keith Kendrick and his colleagues began a series of groundbreaking studies aimed at understanding social communication in sheep (for a review see Tate, Fischer, Leigh, & Kendrick, 2006), and found that their facial processing closely resembled that of humans and monkeys.

Sheep
It is not known whether other non-primate mammals exhibit innate preferences for face-like patterns, or whether their face processing improves with experience. There is a small amount of evidence for visual imprinting in mammals. Newborn voles, which like some birds are mobile and independent soon after birth, will imprint on individual humans and follow them, although it is not known what visual cues are used. However, it is clear that, at least in some non-primate mammals, early experience with faces can significantly shape later behavior. In an unusual experiment, baby goats were raised by mother sheep, and vice versa. As adults, when adopted males of either sex were shown pictures of sheep and goat faces, they preferred to look at females of the foster mother's species (rather than their own sex). Furthermore, when placed in a pen with females of both sexes, they preferentially attempted to mate with the foster mother's sex, demonstrating that early facial experience determines sexual preference.
The authors note that many mammals easily produce and interpret facial expressions. Is this behavior the result of shared and evolutionarily conserved cortical face processing circuits? It is possible that at least some aspects of face perception were inherited from a common ancestor, and not all aspects of face specialization emerged independently in all mammalian superseries. However, it is by no means clear how shared modules for face processing could have been conserved: much of mammalian evolution occurred during the dinosaur era, where most mammals occupied nocturnal niches, thereby reducing the utility of visual cues for social signaling . However, some prominent facial markings such as eye-ring patterns common in extant mammals may have first evolved to make faces more prominent in moonlight, with some extant nocturnal primates being most active. During this nocturnal era, adaptations critical to facial signaling, such as moving ears and elaborate mimetic muscles, became permanent in the mammalian face, although they may have initially had no connection to visual signaling. Following the extinction of the dinosaurs, when some mammals became diurnal, these natural facial patterns and movements may have increasingly served as a basis for personal identification and social signaling.
Clearly, there are many open questions regarding mammalian facial processing and its evolutionary relationship to primate facial processing. Since neither brains nor cognitive processes leave behind a fossil record, the evolutionary history of face perception must be inferred from similarities in behaviors, anatomy, and neural responses between different species. The distinction between homology [homology is the similarity of two organs in creatures that do not belong to the same species] and convergent evolution [convergent evolution is the development of superficially similar characteristics, in creatures of different origins and independently of each other] is particularly difficult.
Homology would require functionally and anatomically similar face-responsive regions to be present in the common ancestor of mammals and primates, a mammal that lived with dinosaurs during the Cretaceous [Cretaceous is the latest period of the Mesozoic era. It lasted about 80 million years, from 145 million years before our time (with the end of the Jurassic period) to 66 million years before our time (with the beginning of the Paleogene period). For some critical aspects of interior processing, this intriguing possibility cannot be ruled out. Alternatively, given that both sheep and macaques are diurnally active and social, their selective cortical mechanisms may have evolved independently, as parallel adaptations of a general visual processing system, driven by similar sources of natural and sexual selection. In any case, the functional and neural similarities of mammalian and primate face perception highlight the possible ancient evolutionary origins of face perception.
Reptiles, birds, and fish lack the sophisticated facial muscles of mammals, so they have a limited repertoire of facial behavior. However, many species do interact socially, with some paying particular attention to the face and head. Like humans, some non-mammalian vertebrates are born with an innate tendency to respond to certain visual patterns. These tendencies can serve as a basis for attraction, or as a signal of danger that causes them to flee. An example of this is in jewel fish, which already 13 days after spawning instinctively avoid pairs of black dots that resemble eyes, even in the absence of previous experience with real eyes. Juvenile and adult fish become very sensitive to this cue and use it to guide their aggressive or avoidant behavior in the dominance hierarchy.

We will also mention here that, interestingly, Dr. Yonat Ashhar recently wrote on the Davidson website that it was demonstrated that the sniper fish is able to recognize human faces. The author cites Kate Newport and her colleagues from the University of Queensland in Australia who studied the snipe (Toxotes chatareus), a freshwater fish that feeds on insects that it hunts by spitting a precise jet of water in their direction. According to her, the researchers used this feature of his and taught him to direct the stream of water to one face and not the other. They trained four fish in this way, which were able to recognize the same face even when they were presented with black-and-white pictures, or ones that showed only the face – and not the whole head. The fish still knew how to distinguish between the faces even without important characteristics such as skin color and head shape. However, it is possible that in the absence of the new cerebral cortex that mammals have [where the fusiform gyrus is found], fish are unable to recognize a more complex stimulus than a face looking directly in their direction.
Newport C, Wallis G, Reshitnyk Y, Siebeck UE. Discrimination of human faces by archerfish (Toxotes chatareus)..Sci Rep. 2016 Jun 7;6:27523.

Sniper fish. Insect fish [The image is taken from the web]
One aspect of avian visual development that is similar to the development of face perception in primates is the innate preference to orient to a particular configural [configuration, spatial shape] stimulus shortly after birth. In some birds, filial imprinting is a behavioral phenomenon in animals in the first stages of their development. This is a unique learning process that is common mainly in birds and poultry, in which the offspring of the animals imitate the behavior of their mother or another authority figure that they were exposed to in a short period of time from the moment they came into the world. Filial – the first generation after the parents – goes through a temporal sequence similar to that seen in humans. For example, chicks are initially attracted to any stimulus with eyes and/or head and neck shape. Then, after a relatively short period of experience, the preference becomes more focused on a single object or a single person, a critical step in filial imprinting. Damage to the latter structure eliminates the filial imprinting. Note that these two sequential orienting mechanisms are similar to those observed in human infants.
The authors point out that the evidence points to social interactions between non-mammalian vertebrates, especially birds, although the role of the face is unclear and probably minor compared to mammals. The eyes appear to be important stimuli for all major vertebrate species, and the ability of corvids to read human gaze is remarkable. Given that birds are so evolutionarily distant from primates and have such different brain structures, it seems likely that their gaze perception mechanisms evolved independently of those of primates. Nevertheless, it is still possible that while their common ancestors, who lived about 320 million years ago, had some sensitivity to the eyes. Ancient circuits in the midbrain, diencephalon, and pallium that mediate this sensitivity may have subsequently evolved into more specialized face-processing circuits in both primates and birds, and possibly many other vertebrates.
Invertebrates
For the sake of completeness, we also address complex visual processing in invertebrates, and ask if there might be any connection to human face perception. Some invertebrates can learn and remember complex shapes and vision is an important social sense for some invertebrate species, including butterflies, and in spiders, horseshoe crabs, eyes appear to be a prominent feature for visually mediated social interaction. While there is good evidence that octopuses can recognize objects, such recognition has not yet been rigorously tested. In cuttlefish, five experiments failed to show vision-based individual recognition, despite rich intrasexual interaction, visual displays and mate guarding.
Some invertebrates can recognize each other visually using "face" cues. In one study, wasps treated familiar people as strangers when artificial markings were added to their faces or abdomens, effectively changing their visual identity. In another, crabs remembered the appearance of their opponents' faces following a direct confrontation. Bees can distinguish and remember human faces, and even use the faces they have learned to recognize new states of these faces. These observations show that relatively simple vision systems are capable of complex differentiation to perform individual recognition.
1] that faces are important to a wide variety of animals, as representative species from all types of vertebrates exhibit behavioral responses to faces that differ from those to non-face stimuli. In primates and sheep, for whom face perception is highly developed and serves as the basis for their social exchanges, the brain contains neurons in many areas dedicated to face analysis. Therefore, for these species, which are the only ones that have been systematically examined, it seems that the behavioral importance of faces fundamentally shaped the processing in the visual pathway.
2] At the most basic level, horizontally paired dark eyes elicit specific behavioral responses from a wide variety of species, including primates, other mammals, birds, reptiles, and fish. For visually oriented animals, the importance of eye detection may be understood in an evolutionary context, as it may indicate the approach of a competitor, predator, or potential mate, all of which have important implications for survival. In addition, some species of primates, sheep and birds have been shown to extract more complex information from faces, such as species or personal identity, gaze direction or level of aggression, with primates showing the most sophisticated face-reading behavior. An intriguing possibility is that the shared perceptual basis reviewed here may be particularly suited to facilitate certain forms of interaction with other species. This can include recognition of threatening species, which is important for survival, as well as playful or nurturing behavior sometimes observed between members of different species living together in unnatural conditions.
3] The authors hypothesize that a pair of dark eyes served as the earliest "face" cue, providing visible markers of sight to all sighted creatures. This important clue may have led to the evolution of selective neural responses in the eyes in animals that lived hundreds of millions of years ago, before the first dinosaur, and perhaps even before the first vertebrates. Other important aspects of face perception, such as the detection of an open mouth or fixed gaze involved in predator detection, may also have very ancient origins and may have further shaped the evolution of visual processing in the brain.
4] The authors hypothesize that the recognition based on faces came later, and probably first concerned the identification of the species and not the individual. Visual species recognition in mammals may have emerged while they were primarily nocturnal, aided by highly visible and stereotypical patterns on the face and head. After the extinction of the dinosaurs, diurnal mammals may have taken advantage of this existing recognition system and adapted it to recognize and remember details based on more subtle cues. For many primates, including humans, face perception takes center stage in social signaling, and the origin of such signaling may lie in the heightened sensitivity to detail afforded by high visual acuity. In addition, the learning of subtle facial expressions and the ability to differentiate between very similar faces require a wide network of brain regions that undergo a long period of experience-dependent development. The dominance of face recognition in primates over that of other animals, demonstrated by the use of configural cues to recognize unfamiliar faces, appears to depend on neural coding that is both systematic and adaptable. Through experiential learning, primates including humans optimize their personal face interpretation mechanism, as both psychophysical and physiological experiments have proven that over time, the brain automatically adjusts itself in light of the variety of faces it experiences.
We will now mention the article by Landi and Fierwald from 2017.
Sofia M Landi & Winrich A Freiwald. Two areas for familiar face recognition in the primate brain. Science/ 2017 Aug 11;357(6351):591-595.
The abstract of the article claims that the very familiarity with matched individuals changes face recognition: familiar faces are recognized more accurately than unfamiliar faces, even under difficult viewing conditions when unfamiliar face recognition fails. The researchers examined the neural basis for this difference. Using whole brain functional magnetic resonance imaging, they found that personally familiar faces engaged the macaque monkeys' face processing network more than unfamiliar faces. Familiar faces also recruited two previously unknown face regions in anatomically conserved locations in the perirhinal cortex and temporal pole. These two areas, but not the core face processing network, responded to familiar faces with a typical non-linear surge of brain activity, similar to the familiar face recognition spiking. In contrast, responses to unfamiliar faces and objects remained linear. Therefore, the two areas of the temporal lobe expand the core of the face processing network into a familiar face recognition system and it is possible that the greater the importance, the more areas are recruited. Hence, as faces become more familiar and more important, additional areas in primates used for identification are apparently recruited.

Another article written by Tomkins and his colleagues is of interest and reports on separate brain areas for human and dog face processing as revealed in awake dogs (Canis familiaris).
Andy M Thompson et al. fMRI Separate brain areas for processing human and dog faces as revealed by awake fMRI in dogs (Canis familiaris)/ Learn Behav. 2018 Dec;46(4):561-573.
The authors begin by saying that functional magnetic resonance imaging (fMRI) has emerged as a viable method for studying the neural processing underlying cognition in awake dogs. In their study, working dogs were shown pictures of dog and human faces. The human faces were of familiar trainers and unfamiliar people and these were also presented with emotional valence (negative, neutral and positive). The dogs' faces were familiar (dog friends) or unfamiliar. The findings revealed adjacent but separate brain regions in the left temporal cortex for human and dog face processing in the dog brain. The human face region and the dog face region were both parametrically characterized by valence indicating that emotion was not the basis for separation. The human face region and the dog face region were not affected by the familiarity theme. Using resting state fMRI data, functional connectivity networks (connectivity fingerprints) were compared and the relevant regions were matched between dogs and humans. Analyzes of these networks found that the human face region mapped to the human fusiform region and the dog face region mapped to the human superior temporal gyrus, two core regions in the human face processing system. The findings provide insight into the evolution of facial processing.
This is very interesting because it can be assumed that their human masters are generally more important to dogs than other dogs. This suggests that the fusiform nucleus is central to recognizing faces that are important to the dog. And we know that this nucleus is central to recognizing individual faces even in humans.
Hence the question arises as to whether it is possible that the fusiform in a variety of relatively higher animals reflects the most important face data processing nucleus among all areas. And is the processing in it done in a better quality, maybe faster and with a higher resolution.
We will add here two aspects related to the fusiiform and shed additional light on its function. One is that with autistics, faces and identifications apparently do not play an important role for them, the human face is not represented there.
Bailey A., Bräutigam S., Jousmaki V, Swithenby S.J.(2005). Abnormal activation of face processing systems at early and intermediate latency in individuals with autism spectrum disorder: A magnetoencephalographic study. European Journal of Neuroscience, 21, 2575–2585.
Secondly, it was demonstrated that for collectors who are devoted to their collections, those collectibles that are of great importance to them are represented in the fusiform region. Something that teaches that this area is central to the visual representation of things that are very important to the individual.
It is therefore said that the facial recognition area and the fusiform in its center play a role in the individual recognition of individuals of the same species and even of other species if these are important and that these recognition abilities developed during evolution especially in social animals.
We therefore hypothesize that the physioform and the facial recognition system that enables individual recognition is the nucleus or core around which the representation of the internalized characters is developing.
In the next conversation we will continue and discuss the work of Olivier Pascalis "The origins of facial processing in humans: phylogeny and ontogeny" and other topics related to facial recognition.
Best regards
Dr. Igor Salganik and Prof. Joseph Levine
Leave a comment