 Prof. Joseph Levine, M.D. is an emeritus associate professor in the Division of Psychiatry, Faculty of Health Sciences, Ben Gurion University in Israel. Prof. Levine is a certified psychiatrist with clinical experience in controlled trials of adult psychiatric disorders and in psychotherapy. He was awarded a NRSAD independent investigator grant for the study of Creatine Monohydrate in psychiatric disorders -- mainly Schizophrenia. He resides and treats patients in Tel Aviv and all of central Israel.
Prof. Joseph Levine, M.D. is an emeritus associate professor in the Division of Psychiatry, Faculty of Health Sciences, Ben Gurion University in Israel. Prof. Levine is a certified psychiatrist with clinical experience in controlled trials of adult psychiatric disorders and in psychotherapy. He was awarded a NRSAD independent investigator grant for the study of Creatine Monohydrate in psychiatric disorders -- mainly Schizophrenia. He resides and treats patients in Tel Aviv and all of central Israel.
Conversation 21: The face recognition center in the brain as a nucleus for creating an internalized character, the social hierarchy, the social brain map and the place of the group leader in the brain
Greetings
In this talk, we will discuss the facial recognition centers in the brain and claim that they are the core or the anchor for creating an internalized character, and we will also present studies that discuss the social brain map of the individual that reflects a social hierarchy, which has possible consequences regarding the hierarchy of internalized characters, and we will say a few words about the leadership of a group and its representation in the brain.
The face recognition centers in the brain.
Face processing researchers often claim that humans spend more time looking at human faces than at any other category of complex visual stimuli. As a result of this unparalleled exposure, adults appear to have an extraordinary level of ability in the area of facial processing. It is this proficiency that is at the center of a debate: Does facial processing have a unique status in the brain?
David J. Kelly and Olivier Pascalis in their review article:
[Olivier Pascalis & David J. Kelly. The Origins of Face Processing in Humans: Phylogeny and Ontogeny. Perspect Psychol Sci. 4: 200-209; 2009.]
It is argued that the answer to this question is an emphatic "yes", and that the evidence supporting this conclusion is overwhelming. They draw on developmental psychology, evolutionary psychology, and comparative psychology to show that face processing in humans and related species is performed by a distributed cognitive system that is unlikely to have developed recently in evolutionary history and is essential to our ability to function as a social species.


Olivier Pascalis—developmental researcher [above]
David J. Kelly – developmental psychologist [below]
Evidence from developmental psychology shows that despite the apparent generality of the face processing system in newborn infants, there are two distinctly specific face abilities found early in life. First, from the age of 2-3 days, babies prefer to see their mother's face when paired with a stranger's face, and secondly, babies show a preference for attractive faces when they are paired with less attractive faces. Both of these preferences can be turned off with some ease. In the case of the mother's face, it is enough to mask the external features (outer contours of the face and hairline) to remove the preference. For attractive faces, the preference can be removed by reversing the stimuli.
In addition to face preference, human infants also demonstrate evidence of categorization—the tendency to group faces into distinct morphological groups. However, infants do not show sensitivity to the subtle differences in morphology that are clearly seen in adults, and instead show a broad preference for "faces" in general. However, face processing abilities develop rapidly from birth, as babies are exposed to more and more faces in their environment. It has been shown that infants begin to show evidence of facial prototype formation at 3 months of age. A prototype is an average face created by encoding several individual faces.
This fact is also supported by the findings that 3-month-old infants show a visual preference for faces that match the gender of their primary caregiver and faces of their own ethnic group. Collectively, these findings demonstrate the importance of experience with faces from the infant's visual environment and how they influence the development of the face processing system.
Evidence from comparative psychology demonstrates that facial processing is not specific to humans, and similarities with other primate species suggest common ancestry. In both human and primate society, facial recognition plays a crucial role. The mature facial systems of primates share several similarities with the human system: for example, the scanning ability of the eyes, personal recognition, and the influence of face inversion effects (faces shown upside down so that the chin is up and the forehead is down). In addition to the similarity in the adult system, the developmental trend is also similar. Faces represent a highly attractive stimulus for primates at an early age, including macaques (Macaca nemestrina), gibbons (Hylobates agilis) and chimpanzees. Although the developmental time scale differs between species, this is true for both general brain development and other cognitive functions. Interestingly, human infants show the ability to recognize primate faces by 9 months of age, demonstrating that the face processing system becomes oriented to human faces during the first year of life. In primates, two-month-old macaques already show a unique preference for their own species. Facial processing is thus highly specialized from an early age in both humans and primates, and may have played an important role in the development of primate societies based on strong and meaningful relationships.

In addition to primates, other social species exhibit similar facial abilities. For example, we mentioned in conversation 20 that sheep show specialized facial recognition abilities. There is even evidence that wasps, which rely primarily on chemical signatures for communication and identification, can also demonstrate hierarchical classification (for example, of a worker) based on facial cues alone. Collectively, these results strongly suggest that a face processing system is not human or even primate-specific, and is unlikely to have been developed recently in evolution.
It is argued that humans and primates have an evolved face processing system that has become specialized as a result of dominant exposure to faces of a single species. According to this interpretation, expert levels of face recognition depend on both a developed mechanism and experience. Therefore, due to the biological irrelevance of various objects in contrast to faces, in studies that attempted to mimic the development of naturally acquired expertise for objects, even intensive exposure from birth would not produce the same "expert" levels of face processing observed in adulthood.
The authors conclude that the ability to individuate is necessary for any species that lives in a social group. Individuation can be accomplished in a variety of ways, and it is likely that most species will use their highly developed senses to accomplish this task. Although primates use their visual abilities, other species may rely on their sense of smell (eg, hamster) or sense of hearing (eg, birds). A general mechanism for individuation may have developed at some point during evolution and is utilized in different ways by different species. Alternatively, such abilities may evolve independently in different species. In any case, successful interaction in social species appears to depend on a highly developed capacity for individual recognition.
The authors suggest that the early face recognition system observed in infants is a combination of evolutionary inheritance, learning in utero and rapid learning after birth. At birth, they note, the representation of the face is crude, and based on the pattern of evolution and is enhanced by proprioceptive acquisition (eg, eye movement). This early template benefits from a fast learning system.
The authors note that the core of the human neural system for face processing as revealed by fMRI studies consists of three cortical regions: the inferior occipital gyrus, the middle fusiform gyrus (fusiform face area; FFA), and the superior temporal sulcus. These areas become more active in humans while viewing human faces rather than other visual objects.
Interestingly, brain imaging using positron emission tomography demonstrated adult-like brain activity in two-month-old babies when looking at faces. However, infant faces also activated areas normally devoted to language in adults, suggesting a possible early connection between the visual and auditory systems or even a more distributed network. It was also found that an area identified as FFA is activated bilaterally by faces in children from the age of 9. A study that compared recognition of objects and faces in children, teenagers and adults, after viewing the objects, similar brain activity was found in all three age groups.
In contrast, FFA activation was found when both older groups were presented with faces, but there were difficulties in isolating the FFA effect in the youngest group. Despite that the findings today remain unequivocal, it can be concluded that the FFA develops from a young age, but the overall pattern of results supports a permanent specialization of the neural network in the face throughout ontogeny [individual development].
Finally, the authors of the article note that facial processing seems to be a complex and multi-faceted task, so we should not expect a simple system to be at its core. Converging evidence from behavioral, electrophysiological, and brain imaging studies provides strong support for a complex neural architecture likely dedicated to performing this task. In addition, comparative studies indicate that faces are very important for various primate species and even other mammals, such as sheep. The studies the authors described in their review indicated that faces may be processed similarly in many species by a common system.
Information about social hierarchy and the structure of the social network.
As mentioned above, we propose that the face perception system [mainly the inferior occipital gyrus, the middle fusiform gyrus (fusiform face area; FFA) and the superior temporal sulcus] are the nucleus for the internalization of the characters, including their attitudes and characteristics, and that these characters build the social self [see previous conversations number 1-3. We note that in addition to this fusiform nucleus there are a variety of other brain structures of importance related to the characteristics of the characters and the relationships between them. In addition, there are various clues to the hierarchy of figures in the brain in humans and even in primates within a social network encoded in the brain. A hierarchy built according to their importance and their kinship and social power. Accordingly, we propose that there is a hierarchy of internalized characters in humans according to their importance within the social self.
We will first discuss the question why is hierarchy important in social systems? What are its advantages? We will mention works describing the map of the social network in the brain and related topics.
Let's first mention the article by Emily Zitek and Taylor Phillips who wrote a review about the cognitive benefits of the existence of a social hierarchy.
Emily M Zitek, L Taylor Phillips, Ease and control: the cognitive benefits of hierarchy. Curr Opin Psychol.; Jun;33:131-135, 2020.
This review identified two cognitive benefits of social hierarchy that may contribute to hierarchy maintenance. First, research suggests that people notice hierarchies automatically, early, and accurately. As a result, hierarchies feel easy to cognitively process, which increases liking and support for the hierarchy. Second, through the clear and predictable structures of hierarchies and the opportunities they provide, hierarchies help people satisfy their need for control, which may lead people to seek and maintain a hierarchy, especially if they hold it in high regard or believe in social leadership. These cognitive benefits of ease of processing and control may have effects on hierarchy performance and people's willingness to change unfair structures.
Another article is by Morrison and his colleagues who first point out that modern human societies exhibit a hierarchical social modularity in which lower order social units such as nuclear families are nested within larger units.
M, Breuer T, Manguette ML, Walsh PD. 2019. Hierarchical social modularity in gorillas. Proc. R. Soc. B 286: 20190681.
It is argued that hierarchical social modularity evolved independently after the chimpanzee-human split. The authors used network modularity analysis and hierarchical clustering to quantify community structure in two communities of lowland gorilla populations.
Both strata closely mapped the social strata found in humans. Genetic data from one population suggested that, as in humans, membership in the social unit was that of kin. The sizes of the gorillas' social units also showed the kind of relative consistency and scale of social stratification observed in humans, baboons, toothed whales, and elephants. These results suggest that the hierarchy of social organization observed in humans evolved much earlier than previously thought and may not simply be a product of social brain evolution unique to the hominid lineage.
Another article is by Thompson and his friends who published an article entitled: A functional brain network architecture that supports the learning of social networks in humans.
Steven H. Thompson, Ari E. Kahn, Emily B. Falk, Jean M. Vettel, Danielle S. Bassett. Functional brain network architecture supporting the learning of social networks in humans: Neuroimage. 2020 April 15; 210: 116498.
The authors note that most humans have the good fortune to live their lives embedded in structured and rich social groups. However, it remains unclear how humans acquire knowledge of these social structures to successfully navigate social relationships. The authors addressed this knowledge gap through interdisciplinary brain imaging research based on advances in network science and statistical learning. Specifically, they collected BOLD MRI data [blood oxygen level-dependent (BOLD) imaging, a technique commonly used to measure brain activity in humans using magnetic resonance imaging (MRI)] while participants learned the community structure of social and non-social networks, to examine whether the learning of these two types of networks was differentially related to the topology of functional brain networks. The authors found that participants learned the community structure of the networks, as evidenced by a slower reaction time when the subject moved between social communities compared to moving within the same community. The study of the community structure of social networks was also characterized by significantly greater functional connectivity of the hippocampus and the temporoparietal junction when moving between communities than when moving within a community. Furthermore, default mode temporoparietal regions were more strongly connected to hippocampal, somatomotor, and visual regions for social networks than for nonsocial networks. Collectively, the results identify neurophysiological foundations of learning in social versus non-social networks, and expand knowledge about the impact of social context on learning processes.
Emily M. Zitek & Larissa Z. Tiedens. The Fluency of Social Hierarchy: The Ease With Which Hierarchical Relationships Are Seen, Remembered, Learned, and Liked. Journal of Personality and Social Psychology, 2012, Vol. 102, no. 1, 98–115.
We will also mention here the research of Emily Zitek and Lisa Tiedens who tested the hypothesis that social hierarchies are social stimuli that are processed more fluently and easily and are therefore more preferred than less hierarchical stimuli.
In the first study, pairs of people who were presented in such a way that a hierarchy was created between them based on their facial dominance were recognized faster than pairs of people who were equal in their facial dominance.
In the second study, a diagram representing hierarchy was recalled more quickly by subjects than a diagram representing equality or a diagram of data comparison. This faster processing led to subjects liking the hierarchy diagram better than the equality diagram.
In a third study, participants were best able to learn a set of relationships they represented hierarchy (asymmetry of power) – relationships where there was an asymmetry of friendship, as compared to relationships where there was symmetry. This led to the fact that relationships represented by the hierarchy were more pleasing to the subjects.
In a fourth study, the participants found it easier to make decisions about a company with a hierarchical structure and concluded that the hierarchical organization has more positive attributes.
Their research also showed that when the nature of social relationships is hard to learn people's preference for hierarchy increases. Together, these results point to one reason why people may tend to prefer hierarchies—because hierarchies are easy to process. Such fluency and ease of processing of social hierarchies may contribute to the construction and maintenance of hierarchies.
We will also mention here the article by Fiske and her friends who claim that hierarchies of power (resources) and status (prestige) are constants that organize human societies.
Susan T. Fiske, Cydney H. Dupree, Gandalf Nicolas, and Jillian K. Swencionis
Princeton University. Status, Power, and Intergroup Relations: The Personal Is the Societal. Curr Opin Psychol. 2016 October; 11: 44–48.
The authors review relevant social psychological literature and identify several converging results concerning power and status. Whether the rank is long-term or temporary, higher ranks create a psychological distance from others, allowing the higher ranks a permanent distance and more accurate treatment of them than the lower ranks. The beliefs that status entails competence are essentially universal. Interpersonal interactions create compensatory trade-offs between providing interpersonal warmth on the one hand and ability and competence on the other. Together with social structures (in which there is persistent inequality), these compromises reinforce beliefs that status goes with ability. Race, class, and gender further illustrate this dynamic. The authors note that although status systems are durable, they can change, and understanding these processes of change is an important direction for future research, as global demographic changes disrupt existing hierarchies.
Van Berkel and her friends claim that "hierarchy may have a psychological advantage over equality in that it is familiar, repetitive, and socially effective."
Laura Van Berkel, et al. Hierarchy, Dominance, and Deliberation: Egalitarian Values Require Mental Effort. Volume 41, Issue 9.
These authors show that when the participants' deliberate thinking was limited – by an increase in the blood alcohol level, which creates a cognitive load that makes it difficult to think without considerable effort, or to react quickly – the participants increased their support for the hierarchy. Accumulating evidence such as this suggests that group power and status distinctions are relatively automatic and spontaneous, perhaps even the human default. Hence, according to the authors, ranking power and status are not going to disappear anytime soon.
Farrow and his colleagues claim that the perception and judgment of social hierarchies are an integral part of social consciousness and that hierarchical judgments can be self-referential or allocentric (concerning two or more external agents).
Farrow, T.F.D., Jones, S.C., Kaylor-Hughes, C.J., Wilkinson, I.D., Woodruff, P.W.R., Hunter, M.D., Spence, S.A. (2011) Higher or lower? The functional anatomy of perceived allocentric social hierarchies, NeuroImage, 57 (4), pp. 1552-1560.
The authors sought to investigate the brain regions involved in judging allocentric social hierarchies. 22 healthy male subjects underwent three brain imaging [fMRI] scans. During scanning, subjects answered questions concerning visually presented target pairs about the relative superiority of the human individual within a particular social hierarchy or about their perceived degree of social alliance (ie, whether they were "friends or foes"). Subjects also made judgments regarding the age, gender, and status of the target couples to control for confounding factors and performed a basic numerical task. The response times increased according to the increasing load of the processing. Both social hierarchy and social alliance judgments activated the left ventral prefrontal cortex (VLPFC), the left dorsal inferior frontal gyrus (IFG), and the bilateral fusiform gyri. In addition, social alliance judgments activated right dorsal IFG and medial prefrontal cortex. Compared directly to social alliance, social hierarchy judgments activated the left orbitofrontal cortex. Recognizing the presence of social hierarchies and judging the relative status of others within them implies activity of the cognitive brain executive area, and specifically the VLPFC. The researchers' findings are relevant to 'normal' social cognition performance, but their method also provides a means to study the dissocial brain in personality disorder and schizophrenia disorder where executive brain function may be impaired.
We will continue and mention here Courtney and Mayer who wrote an interesting article about how the representation of the self vis-a-vis others in the social brain reflects a social relationship.
Andrea L Courtney & Meghan L Meyer ; Self-Other Representation in the Social Brain Reflects Social Connection. J Neurosci. 2020 Jul 15;40(29):5616-5627.

Megan Mayer – researcher of the social brain.
These authors first note that social connection is critical to well-being, but how the brain reflects our attachment to other people remains largely unknown. The authors' group combined univariate and multivariate brain imaging analyzes to assess whether and how the brain organizes representations of others based on how closely they relate to our own identity. During brain imaging [fMRI scanning] 43 female and male human participants completed tasks involving self, five close others, five acquaintances, and five celebrities. The authors examined neural responses to self and others in the area of the brain associated with self-representation (the prefrontal cortex: mPFC) and in addition in the whole brain. Their study shows that the structure of self-versus-other representation in the mPFC and throughout the social brain appears to group targets into three social categories: self, members of social networks (including other relatives and acquaintances), and celebrities. Furthermore, both univariate activation in the mPFC and multivariate self-similarity in the mPFC and across the social brain increased with subjective ratings of self-other closeness. Critically, participants who were less socially connected (ie, more lonely) showed changes in self-mapping in social brain regions. In the mPFC primarily, loneliness was associated with reduced representational similarity between self and others. The social brain likely stores information about broad social categories as well as proximity to the self. Furthermore, these results suggest the possibility that feelings of chronic social detachment may be reflected by a more “lonely” neural self-representation. According to the authors, this means that social connection is critical to well-being, but it is still not entirely clear how the brain reflects our connection to people. The authors found that the social brain stratifies neural representations of people based on our subjective connection to them, and groups separately people who are in our social network and those who are not. Furthermore, the people we feel closest to are represented in the closest way to ourselves.
Finally, lonelier people also appear to have a "lonelier" neural self-representation in the mPFC, as loneliness weakened the similarity between self and other neural representations in this region. The social brain appears to hierarchically map our interpersonal connections, and changes in this hierarchical map may help explain why single people endorse statements such as "people are around me but not with me."
We will point out that this article is particularly interesting for our hypothesis regarding the existence of a "jury" or "board" of internalized figures significant to us that build our social self, within which there is a hierarchy.
Tavares and her colleagues who studied such a social brain map first point out that deciphering the neural mechanisms of social behavior drove the growth of social neuroscience. The precise computations of the social brain, however, remain elusive.
Tavares RM, Mendelsohn A, Grossman Y, Williams CH, Shapiro M, Trope Y, Schiller D. A Map for Social Navigation in the Human Brain. Neuron. 2015 Jul 1;87(1):231-43.
These authors investigated how the human brain tracks ongoing changes in social relationships using functional brain imaging. Participants were main characters in a role-playing game where they were supposed to find a new home and job through interactions with virtual cartoon characters. The authors found that a two-dimensional geometric model of social relations, a "social space" framed by power and affiliation, predicted hippocampal activity. Furthermore, participants who reported better social skills showed stronger covariance between hippocampal activity and "movement" through "social space." The results indicate that the hippocampus is essential for social cognition, and imply that beyond framing physical locations, the hippocampus computes a more general, inclusive, abstract, and multidimensional cognitive map consistent with its role in episodic memory. We will point out that the existence of such a map representing a "social space" framed by power and affiliation first builds a hierarchy of affiliation that is proximity and secondly a hierarchy of power relations.
We will also note that the family of each of us exists for the most part in significant proximity to us.
Steven Flatek and Shelly Kemp asked if the family has a special representation in the brain? And accordingly, they conducted an event-related brain imaging [fMRI] study about recognizing familiar faces, family faces, and the self.
Steven M Platek, Shelly M Kemp. Is family special to the brain? An event-related fMRI study of familiar, familial, and self-face recognition. J. Neuropsychologia. 2009 Feb;47(3):849-58; 2008.

Steven Platek—engages in research psychology.

Shelly Kemp – research psychology.
The authors note that the face processing network has evolved to respond differentially to different classes of faces according to their relevance to the perceiver [essentially arguing about the existence of a hierarchy based on relevance to the person]. For example, self, familiar and unfamiliar faces are associated with the activation of different neural substrates. Family should represent a special type of face stimuli with high relevance to individuals, because misidentification of kinship can have severe consequences (e.g., incest, infidelity). Therefore evolution should have favored specific mechanisms for recognizing relatives.The authors used fMRI [functional brain imaging] to investigate the neural substrates associated with viewing relatives' faces compared to other types of faces (e.g., own faces, familiar faces, and unfamiliar faces), and examined the extent to which self-face similarity activated similar neural substrates . Familiar faces, in contrast to unfamiliar faces, activated brain substrates related to substrates that work for self-face recognition. Comparing close faces to the faces of friends activated the brain regions called the posterior cingulate and the cones. Similar medial posterior brain substrates were recruited when contrasting self-like faces with distorted faces of relatives, suggesting that these regions may represent computational processing regarding face familiarity and identity. On the other hand, discrimination of self-similar faces following familiar face transformation activated anterior medial substrates (anterior cingulate gyrus [ACC] and medial prefrontal cortex [mPFC]). These findings, and region-of-interest (ROI) analysis, highlight the role of the extended face processing network in distinguishing kin from familiar non-kin members of one's social group. That is, a hierarchy of 1] the self, 2] relatives, 3] familiar friends who are not relatives.
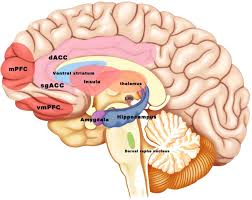
[Looking at the inside of the right hemisphere of the brain]
Anterior cingulate gyrus [ACC] and medial prefrontal cortex [mPFC] and see also location of amygdala and hippocampus].
Carolyn Zink and her colleagues conducted a brain imaging [fMRI] experiment in which two groups participated in two social hierarchy experiments [a stable hierarchy versus an unstable hierarchy in which the position in the hierarchy may change], alongside an experiment in a control group of the non-social hierarchy. In this investigation, the authors used skill differences and associated rank symbols to create a hierarchy.
In their study, the authors identified striking differential neural responses based on status when viewing another person. Position in a hierarchy can be fixed or changeable, and this aspect of social stratification has significant implications for individuals. In nonhuman and human primates, the more subordinate position in stable social hierarchies is associated with greater stress, whereas in dynamic hierarchies, the dominant position experiences the most stressors due to increased competition from others and instability during periods of reorganization, and may be associated with greater health risk. In order to investigate differences in brain representation between stable and unstable hierarchical information processing, the authors tested the stability of the hierarchy in two experiments. It is important to note that in addition to the stability of the hierarchy, the authors also tested social specificity, through a non-social control experiment, which allows separation between the neural processing of general hierarchical information (that is, one that is ranked relative to an inanimate being) and social hierarchical information (that is, ranked relative to other humans).
Across all hierarchical settings (stable, unstable, and antisocial), brain activity was significantly greater in the occipital/parietal cortex, ventral striatum, and parahippocampal cortex, suggesting that these brain regions are active in the neural encoding of hierarchical rank, regardless of stability or social nature specific to the hierarchy.
While these regions did not appear to differentiate between social and nonsocial hierarchical information, activation of the dorsolateral prefrontal cortex [DLPFC] was only seen in a social context, suggesting that DLPFC involvement in hierarchical information processing is specifically social. The authors' data support the idea put forward earlier that the DLPFC plays a role in interpersonal judgment, including the assessment of social status. Furthermore, the DLPFC has been implicated in compliance with a social norm, a process strongly influenced by perceived social hierarchical rank. Consistent with the resulting social specificity of DLPFC activity, it has been reported that the role of the DLPFC in social norm compliance was significantly more pronounced in a social context compared to a non-social context.
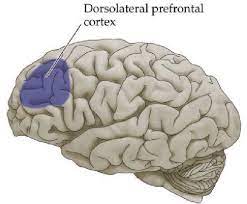
Dorsal lateral cerebral cortex in the prefrontal brain region.
Interestingly, the social unstable hierarchical setting evoked multiple neural responses that were not produced with the other (stable and non-social) hierarchical settings. The unstable social hierarchy resulted in an increase in activity in many areas related to social emotional processing and social cognition. The amygdala, in particular, has been reported to be involved in the processing of social emotional stimuli, as well as in social anxiety related to hierarchical challenge. In addition, it was taught that activity in the thalamus, amygdala and posterior cingulate is regulated by social emotional stimuli. The authors found significant positive correlations between the activity in those areas and the level of positive affect of subjects in the upper hierarchical position. The authors concluded that the activity in these areas represented an emotional arousal response for subjects in a higher hierarchical position that arises only when the hierarchy is dynamic; that is, when the subject's performance can have social hierarchical consequences.
The MPFC [medial prefrontal cortex], known to play a central role in social cognition, was also uniquely activated within the unstable social hierarchy when the subjects viewed a person higher in the hierarchy versus someone inferior to them. The MPFC is particularly associated with recognizing the intentions and motives of other people (mentalization) and forming judgments of other people (person perception), including how others see us (reputation). There is evidence that the MPFC is involved in forming impressions only in the social domain (ie judging people versus inanimate objects), a claim supported by a variety of data.
The data delineating brain regions that are uniquely activated by a subject higher in the hierarchy compared to the one lower than him in the unstable hierarchical environment are in good agreement with the role of hierarchical rank in Festinger's social comparison theory (Festinger, 1954). Wheeler proved that humans prefer to make upward comparisons, that is, comparisons with superior others, which provokes a negative effect, and that this tendency is stronger with higher levels of motivation to improve (Wheeler, 1966), as is possible only in the unstable hierarchical environment. In fact, there is a paradox arising from this: people with the greatest desire for success have the greatest tendency to make social comparisons with superior others, which leads to negative emotions (Wheeler, 1966).
In conclusion, the above study provides a characterization of the neural correlates associated with the processing of social hierarchies in humans. In this initial investigation, the authors used skill differences and associated rank symbols to create a hierarchy. The findings show that brain responses to superiority and inferiority in a social hierarchy are separate [dissociative], even in the absence of explicit competition, both when encountering a person of a certain status and when faced with an outcome that can affect the person's current position in the hierarchy.
The leader at the top of the hierarchy.
Finally, Tingting Zhou and others interestingly demonstrated that increasing the activity of certain brain cells can help a mouse climb the social ladder and even to the top.
Tingting Zou et al. History of winning remodels thalamo-PFC circuit to reinforce social dominance. Science; Vol 357, Issue 6347; pp. 162-168, 2017.
Neurons in an area called the dorsomedial prefrontal cortex appear to control whether male mice are dominant or submissive to other males, these researchers report in the journal Science. The finding adds to previous evidence that this brain region is involved in social interactions in mammals.
As a rule, male mice compete to establish a social hierarchical order. When each mouse knows its place, there is usually less social conflict in the long run. And as a rule mental strength and a history of victory play an important role in determining social dominance. However, the neural circuits mediating these intrinsic and extrinsic factors remain unclear. Working in mice, the authors identified a neuronal population of the dorsomedial prefrontal cortex (dmPFC) whose activation or inhibition causes victory in a competition with another mouse and an increase in the hierarchy or, conversely, a loss and a decrease in the hierarchy.
Another very interesting work that suggests that certain brain characteristics are related to leadership is by Mayer and his colleagues.
Meyer CJ, Cassidy KA, Stahler EE, Brandell EE, Anton CB, Stahler DR, Smith DW. Parasitic infection increases risk-taking in a social, intermediate host carnivore. Commun Biol. 2022 Nov 24;5(1):1180.
The authors note that Toxoplasma gondii is a protozoan parasite capable of infecting any warm-blooded species and can increase risk-taking behavior in an animal host by affecting the living brain. Despite extensive laboratory research on the effects of T. gondii infection on behavior, very little is understood about the effects of toxoplasmosis on wild intermediate host behavior. Yellowstone National Park, Wyoming, USA, has a diverse carnivore community that includes gray wolves (Canis lupus) and cougars (Puma concolor), intermediate and definitive hosts of T. gondii, respectively. Here, the authors used 26 years of behavioral, spatial, and serological data from wolves to show that wolf territory overlapping areas of high cougar density was an important predictor of infection. In addition, seropositive wolves were more likely to make high-risk decisions such as dispersing the pack and becoming pack leaders, two factors critical to individual survival and birth rates essential to wolves.

Due to the social hierarchy within the wolf pack, the authors hypothesize that the behavioral effects of toxoplasmosis may create a feedback loop that increases spatial overlap and transmission of the disease between wolves. These findings show that parasites have important consequences for intermediate hosts, beyond acute infections, through behavioral effects. Especially in a social species, these effects can leap beyond individuals and affect groups, populations and even ecosystem processes.
From a different direction in relation to leadership, Psychogios and Dimitriadis point out that relationship-based approaches to leadership represent one of the fastest growing areas of leadership and help us understand how to create better organizational leadership.
Psychogios A, Dimitriadis N. Brain-Adjusted Relational Leadership: A Socially Constructed Consciousness Approach to Leader-Follower Interaction. Front Psychol. 2021 Jul 13;12:672217.
The authors note that relationship-based approaches emphasize the relationship and interaction between the leader and the follower. The emphasis is placed on how they communicate and influence each other in achieving mutual goals. Leaders are known to be related to followers and vice versa in the sense of meeting the needs of others towards achieving mutual goals. Leaders and followers are an essential part of this social process, implying that they lose their traditional identity rooted in the formal organizational structure (manager-subordinate) and become inseparable players in the process of co-constructing leadership. What is less known is how leadership actors relate to each other and especially how they try to figure out how to do this in workplaces.
What is even less understood is the importance and role of consciousness in this relationship. What's more, it seems that consciousness is both a basic element and a very elusive element in human relations. Therefore, this paper conceptually examines the concept of consciousness in the context of the social theory of mind to argue that leadership actors need to rethink their approach to individuality and instead focus on interdependent relationships with one another. This article contributes to the field of neuromanagement by introducing the concept of homo relationis. In this respect, we propose that leadership is not only a socially constructed element but also a social brain phenomenon that requires an understanding of the human brain as a social organ.
And again from a different angle,Vanzella and his colleages examine the subject of the leader or leader, this time in a musical composition:
Vanzella P, Ballardin JB, Furucho RA, Zimeo Morais GA, Braun Janzen T, Sammler D, Sato JR. fNIRS Responses in Professional Violinists While Playing Duets: Evidence for Distinct Leader and Follower Roles at the Brain Level. Front Psychol. 2019 Feb 5;10:164.
According to them, music played in ensembles is a naturalistic model for studying joint action and leader-follower relationships. Recently, the investigation of the brain foundations of joint musical actions has received attention; however, the brain correlates underlying the leader and follower roles in music performance remain elusive. The present study addressed this question by simultaneously measuring the hemodynamic correlates of the functional neural activity evoked during the performance of a naturalistic violin duet using fNIRS.

The findings revealed different patterns of functional brain activation when musicians played violin 2 (follower) than violin 1 (leader) part in duets, both compared to solo performance. More specifically, the results indicated that musicians who played violin part 2 [the follower] had greater oxy-Hb [oxy means oxygen, Hb means hemoglobin] activation in temporoparietal (p = 0.02) and somatomotor (p = 0.04) regions. During duet mode relative to solo. On the other hand, there were no significant differences in the activation of these areas between the duet/solo conditions during the performance of the part of violin 1 (leading, p's > 0.05). These findings suggest that ensemble cohesion during a musical performance may impose certain demands when musicians play in the follower mode, particularly in brain regions associated with dynamic social information processing and motor simulation. This study is the first to use hyperscanning fNIRS technology to simultaneously measure the brain activity of two musicians during a performance of a naturalistic music ensemble, and opens new avenues for the study of the brain correlation underlying joint musical actions with multiple subjects in a naturalistic environment.
To summarize, we have emphasized here the importance of hierarchy, its brain representation, and its evolutionary origins. The importance of the social brain map and evidence of brain representation for leadership.
If you look at the previous conversations you can see that first we claim that the group of internalized characters acts according to ways of acting of external groups in real environment. Secondly, we support that the internalized characters who build the social self demonstrate social hierarchy in different ways and often have a leader [sometimes even more than one] which we called the "inner dictator" or “dictator-self” [tyrant in the sense of a leader who has the ability to censor and influence other internalized characters]. From this it may be possible to claim that the data above do not contradict and perhaps even partially and indirectly support our hypotheses [see previous conversations].
Yours until the next talk,
Dr. Igor Salganik and Prof. Joseph Levine.
Leave a comment